| Sittaco/Динозавры | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд: платеозавр Plateosaurus engelhardti; стиракозавр Styracosaurus albertensis (справа) и Scolosaurus cutleri. | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Динозавры | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Dinosauria Owen, 1842 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| Основные клады | |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| Распространение ископаемых остатков мезозойских динозавров | |||||||||||||||||||||||||||||||||||||||||
 | |||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 233,23 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
Диноза́вры[3] (лат. Dinosauria) — клада диапсидных рептилий[a] из группы архозавров. Динозавры возникли в среднем или позднем триасе, в промежутке времени между 245 и 233,23 млн лет назад. Они стали доминирующими наземными позвоночными после триасово-юрского вымирания 201 млн лет назад, однако большинство их основных групп не пережили мел-палеогеновое вымирание 66 млн лет назад. В конце юрского периода[b] внутри тероподовых динозавров возникли птицы, которые стали единственной группой динозавров, пережившей мел-палеогеновое вымирание. Условно всех динозавров, за исключением птиц, называют нептичьими динозаврами[8].
Динозавры — разнообразная группа с таксономической, морфологической и экологической точек зрения. Палеонтологи описали по ископаемым остаткам более 900 различных родов и 1000 видов нептичьих динозавров. Динозавры представлены на каждом континенте, как современными видами (птицами), так и ископаемыми. Птицы, насчитывающие более 10 800 современных видов, являются одной из самых разнообразных групп позвоночных. Бо́льшая часть исследований, проведённых с 1970-х годов, показала, что нептичьи динозавры, как и птицы, были активными животными с повышенным обменом веществ и многочисленными приспособлениями к социальным взаимодействиям. Динозавры занимали многочисленные экологические ниши и освоили разнообразные типы питания. Все динозавры — яйцекладущие животные, причём способность к строительству гнёзд разделяется как птицами, так и многими нептичьими динозаврами.
Хотя предки динозавров передвигались на двух конечностях, многие вымершие группы динозавров включали в себя передвигавшиеся на четырёх конечностях виды, а некоторые из этих видов были способны как к передвижению на двух конечностях, так и на четырёх. Сложные демонстрационные структуры, такие как рога и гребни, были широко распространены среди динозавров, а у некоторых из них имелись костяная броня и шипы. В то время как современные птицы, как правило, невелики из-за ограничений размера, связанных с их способностью к полёту, многие доисторические динозавры, в том числе и птицы, достигали больших размеров — рекордсменами в этом плане являлись завроподы группы титанозавров, среди которых были крупнейшие когда-либо существовавшие наземные животные. Тем не менее, представление о том, что все нептичьи динозавры обладали гигантскими размерами, является ошибочным; многие из них были сравнительно небольшими.
С тех пор, как первые окаменелые кости динозавров были опознаны и признаны в начале XIX века, их ископаемые скелеты стали главными достопримечательностями музеев по всему миру, а динозавры превратились в неотъемлемую часть популярной культуры. Большие размеры многих нептичьих динозавров, а также распространившийся чудовищный и фантастический образ, обеспечили их регулярное появление в бестселлерах и фильмах. Неугасающий общественный интерес к этим животным привёл к значительному финансированию палеонтологии и постоянному освещению новых открытий в средствах массовой информации.
История изучения
Первые находки

Ископаемые остатки динозавров были известны ещё на ранних этапах развития человечества, но их природа долгое время оставалась неустановленной. До того как было доказано существование процесса фоссилизации (окаменения), ископаемые кости часто считались только похожими на настоящие и, соответственно, не воспринимались как некогда принадлежавшие живым существам[9][10]. Однако ископаемые следы динозавров и других архозавров уже привлекали внимание разных народностей мира[10].
К 1600-м годам были изучены некоторые геологические процессы, но учёным ещё предстояло осознать явление вымирания видов[9]. К этому времени относятся ранние попытки научного описания окаменелостей динозавров, в том числе описание оксфордским профессором Робертом Плотом[англ.] в его работе «Естественная история Оксфордшира» (1677) бедренной кости, вероятно, принадлежавшей мегалозавру. Сам Плот верно определил кость как бедренную, но пришёл к выводу, что кость, скорее всего, принадлежала человеку, возможно, титану[11].
Начало последовательного изучения

В конце XVIII и начале XIX века французский натуралист Жорж Кювье (1769—1832) предоставил доказательства существования полностью вымерших видов. Он обобщил накопленные ранее материалы, основав сравнительную анатомию как самостоятельную биологическую дисциплину о сходствах и различиях в анатомии разных таксонов, а также базирующуюся на принципах сравнительной анатомии палеонтологию в её современном виде, науку, объектом изучения которой являются ископаемые остатки организмов прошлого[12][13].
В XIX веке сравнительно-анатомический подход впервые был применён к окаменелостям динозавров, которые теперь описывались с точки зрения биологической систематики[9]. В 1824 году президент Королевского геологического общества Уильям Баклэнд выступил с докладом о находке, сделанной в 1815 году в юрских сланцах Стоунзфилда (графство Оксфордшир) и состоящей из нескольких костей «допотопного» животного. Прибегнув к помощи Кювье, Баклэнд классифицировал находку как ископаемые остатки гигантской хищной ящерицы (лат. Sauria) и, соответственно, назвал её мегалозавром — «огромным ящером»[14][15]. В 1826 году Гидеон Мантелл, хирург из Льюиса (графство Суссекс), действительный член Линнеевского общества, аналогичным образом представил в Геологическом обществе найденные им зубы ранее неизвестного вида, которому он дал название игуанодон (букв. «игуанозубый») за сходство этих зубов с зубами игуан[16][17]. Он же в 1833 году описал гилеозаврa — представителя анкилозавров[18].

В 1842 году английский биолог сэр Ричард Оуэн констатировал несомненное сходство между этими тремя видами: все они были сухопутными, держали конечности под телом, обладали характерным строением таза и отличались своими гигантскими размерами от всех современных рептилий. Оуэн выделил для этих животных особый подотряд ящериц, назвав его Dinosauria[19][9]. Название таксона происходит от др.-греч. δεινός (deinos) — «страшно великий» и σαῦρος, σαύρα — «ящер, ящерица»[19][20]. Хотя таксономическое название динозавров часто интерпретируется как некое указание на их зубы, когти и другие внушающие страх особенности и поэтому ошибочно переводится как «ужасные ящеры», Оуэн подразумевал размер и величие этих животных[21]. Советский палеонтолог И. А. Ефремов использовал для обозначения таксона Dinosauria русскоязычное название «дивоящеры», представляющее собой кальку с латинского[22]. Кроме того, следует учитывать, что в настоящее время динозавры больше не классифицируются как ящерицы, а большой размер, впечатливший Оуэна, хотя и был характерен для многих динозавров, не разделялся всеми представителями этой группы[9].
Открытие в 1858 году в США хорошо сохранившегося скелета гадрозавра опрокинуло представление о динозаврах как о четвероногих животных, показав, что динозавры могли ходить на двух ногах (хотя конкретно гадрозавр оказался четвероногим). В последующие несколько десятилетий были открыты представители большинства основных групп динозавров; важная заслуга в этом принадлежит американским палеонтологам Гофониилу Маршу и Эдварду Копу, которые открыли и описали в общей сложности 142 новых вида, включая апатозавра, бронтозавра, диплодока, стегозавра, трицератопса и других. Накопление материала привело к тому, что в 1887 году британский палеонтолог Гарри Сили разделил динозавров на отряды птицетазовых и ящеротазовых, но при этом сочёл сам таксон Dinosauria относящимся к искусственной группе, из-за чего он на долгое время почти вышел из употребления в научных кругах.
Ренессанс динозавров

В рамках первой половины двадцатого века бо́льшая часть научного сообщества ошибочно полагала, что динозавры были громоздкими, вялыми животными. В 1964 году находка дейнониха произвела новую научную революцию, так как по строению динозавра было ясно, что он передвигался относительно быстро, из чего следовал вывод, что он был теплокровным. Идея теплокровности заставляла пересмотреть старые представления не только о физиологии, но и о поведении динозавров, чему появились подтверждения в 1979 году, когда были получены доказательства родительского инстинкта и социального поведения майазавры (высиживание, защита и выкармливание детёнышей). Наконец, сравнение верхних конечностей дейнониха с крылом птицы заставило предполагать их близость и происхождение птиц от динозавров, а затем и принадлежность птиц к этой группе, чему ещё одним доказательством стало открытие следов оперения у ряда нептичьих динозавров. Таким образом, большинство исследований, проводимых с 1970 года, указало на то, что динозавры были активными животными с повышенным метаболизмом и способными к различным типам социального взаимодействия. Метод филогенетического анализа позволил доказать монофилию динозавров и снова выделить их в качестве полноценного таксона.
Классификация
Систематическое положение

В настоящее время систематика динозавров обычно рассматривается с точки зрения кладистики, метода классификации, требующего, чтобы все таксоны (клады) были монофилетическими, то есть чтобы они включали всех потомков общего предка, что позволяет строить классификацию исключительно по объективному критерию родства. До 1970-х годов наибольшим влиянием обладала эволюционная таксономия, отличающаяся от строгой кладистики ранжированием таксонов и выделением парафилетических таксонов, то есть таксонов, не включающих всех потомков общего предка; данный подход отвергается подавляющим большинством современных палеонтологов позвоночных как слишком субъективный и не приспособленный к отображению истинных эволюционных взаимоотношений[23][24].
Важным следствием применения кладистики в классификации динозавров является то, что птицы оказываются не потомками или родственниками динозавров, как в эволюционной таксономии, а непосредственно их подгруппой, поскольку они попадают под все существующие филогенетические определения клады Dinosauria[8][24][25][26][27][28]. Филогенетически птицы глубоко вложены в состав группы динозавров-теропод (Theropoda), в пределах которой относятся к целурозаврам (Coelourosauria), а именно к манирапторам (Maniraptora)[29][30]. По мере обнаружения большего количества ископаемых остатков теропод, в той или иной степени родственных современным птицам, граница между «птицами» и «не-птицами» становится всё более размытой[27].

В кладистике динозавры, в том числе и птицы, научно классифицируются как представители клады или класса рептилий (Reptilia)[32][33][34], также известной под лишённым культурных предубеждений названием «завропсиды» (Sauropsida). Одной из подгрупп рептилий, или завропсид, являются архозавры (Archosauria), которые, в свою очередь, подразделяются на две ветви: псевдозухии (Pseudosuchia), куда относятся крокодилы и их вымершие родственники («крокодилья» ветвь), и авеметатарзалий (Avemetatarsalia), куда относятся птицы и их вымершие родственники («птичья» ветвь)[35]. Ближайшими ныне живущими родственниками нептичьих динозавров, птерозавров (летающих ящеров) и некоторых менее разнообразных таксонов являются птицы и, соответственно, все они являются представителями авеметатарзалий. Нептичьи динозавры стоят ближе к птицам, чем к современным представителям традиционного класса пресмыкающихся, не только филогенетически (эволюционно), но и по многим особенностям своей анатомии и физиологии; тем не менее, уровень интеллекта большинства из них, вероятно, был типично рептильным.
Разнообразие
Динозавры являются чрезвычайно разнообразной кладой: по состоянию на 2018 год достоверно идентифицировано более 900 родов нептичьих динозавров и, по данным на 2016 год, 1124 вида[36][37]. В 2016 году предполагаемое количество видов динозавров, существовавших в мезозойскую эру, было оценено в 1543—2468[37][38]. По оценке 2006 года, общее число родов, сохранившихся в палеонтологической летописи могло достигать около 1850, около 75 % из которых ещё не обнаружено[39]. В более раннем исследовании, опубликованном в 1995 году, предполагается, что существовало около 3 400 родов нептичьих динозавров, из которых многие не сохранились в палеонтологической летописи[40]. По данным Международного союза орнитологов, на январь 2021 года известно 10 806 современных видов птиц[41].

Динозавров в целом можно описать как архозавров с задними конечностями, удерживаемыми прямо под телом[англ.][42]. Многие группы доисторических животных, такие как ихтиозавры, мозазавры, плезиозавры, птерозавры и ранние синапсиды, такие как диметродон, неспециалистами часто ошибочно воспринимаются как динозавры, хотя научно не классифицируются как представители этой группы[43][44]. Ни одно из перечисленных животных не обладало выпрямленными задними конечностями, присущими настоящим динозаврам[45]. Положение конечностей динозавров обусловлено развитием углубления в области таза, обращённого вбок (обычно открытого углубления и соответствующей обращённой внутрь головки на бедре[30]). Их вертикальное положение позволило динозаврам при движении легко дышать и, вероятно, повышало уровень выносливости и активности. По этим параметрам они превзошли ползающих рептилий[46]. Вертикальные конечности, вероятно, также помогали поддержать развитие большого размера, уменьшая изгибающие напряжения на конечностях. У некоторых не относящихся к динозаврам архозавров, включая равизухий, также имели вертикальные конечности, но добились этого путём изменения строения тазобедренного сустава[47].
Динозавры были доминирующими наземными позвоночными юрского и мелового периодов. Остальные группы животных, за исключением некоторых других архозавров, в особенности наземных крокодиломорф, в основном были сильно ограничены в своих размерах и занимаемых ими экологических нишах. Млекопитающие, к примеру, редко превышали размера современной кошки, и нередко становились добычей плотоядных динозавров[48]. Заметным исключением является Repenomamus giganticus массой 12—14 кг, который, насколько известно, питался в том числе и мелкими динозаврами, такими как детёныши пситтакозавров[49].
Определение и подгруппы

Согласно формальному филогенетическому определению, рекомендуемому ФилоКодом[англ.]*, таксон Dinosauria соответствует наименьшей кладе, включающей Iguanodon bernissartensis Boulenger in Beneden, 1881 (Ornithischia), Megalosaurus bucklandii Mantell, 1827 (Theropoda) и Cetiosaurus oxoniensis Phillips, 1871 (Sauropodomorpha)[1][50]. Ранее было также дано несколько других определений, в частности, не учитывающие возможного близкого родства между тероподами и птицетазовыми, где вместо двух ящеротазовых указывается только одно (Megalosaurus bucklandii, Passer domesticus или веерохвостые птицы); вместо Iguanodon bernissartensis может также указываться Triceratops horridus. У М. Барона и коллег (2017) таксон Dinosauria определяется как наиболее инклюзивная клада, включающая Triceratops horridus, Passer domesticus и Diplodocus carnegii.
В составе динозавров выделяется три основных хорошо морфологически обособленных группы (см. таблицу «Основные подгруппы динозавров»): тероподы (Theropoda), завроподоморфы (Sauropodomorpha) и птицетазовые (Ornithischia). Первые две обычно относят к кладе ящеротазовых, или заврисхий (Saurischia); к ящеротазовым также могут относиться некоторые базальные (ранние) таксоны, более близкие к тероподам и завроподоморфам, чем к птицетазовым (например, Herrerasauria). По такой системе, все динозавры относятся либо к птицетазовым, либо к ящеротазовым. Анатомически ящеротазовые и птицетазовые наиболее заметно различаются в строении таза. Тем не менее, их названия носят несколько условный характер, так как птицы произошли от более ранних представителей ящеротазовых и, соответственно, по принципам кладистической классификации относятся именно к ящеротазовым, а не к птицетазовым[51]. Эта путаница имеет историческую основу: в 1887 году, когда Гарри Сили впервые разделил динозавров на птицетазовых и ящеротазовых, происхождение птиц от динозавров и, тем более, конкретно от ящеротазовых, было не более чем спорной гипотезой, а о включении их в состав этого таксона тогда не шло и речи[52][53].
У большинства нептичьих ящеротазовых динозавров, как и у их предков, подвздошные кости направлены краниально или вперёд, где они соединяются с крестцом, седалищные — вниз и назад, а лобковые направлены вперёд и вниз. Эта основная форма была модифицирована поворотом лобковой кости назад в различной степени у представителей некоторых таксонов (герреразавр[54], теризинозавроиды[55], дромеозавриды[56] и птицы[27]). У птицетазовых динозавров седалищные и подвздошные кости устроены также, а лобковые кости имеют две ветви, направленные в разные стороны: одна — вперёд, а вторая — назад, параллельно седалищным костям[57].
Альтернативная классификация была предложена М. Бароном и коллегами в 2017 году. По их мнению, тероподы более родственны птицетазовым, чем завроподоморфам, и могут быть объединены с первыми в кладу орнитосцелид (Ornithoscelida). Хотя гипотеза Барона и коллег была поддержана некоторыми отдельными авторами, в настоящее время она остаётся спорной, равно как и филогенетические взаимоотношения между основными группами динозавров.
| Основная клада | Типичное строение таза | Крупная подгруппа | Краткая харатеристика | Некоторые представители |
|---|---|---|---|---|
Ящеротазовые (Saurischia)
| 
| † Herrerasauria † Герреразавриды (Herrerasauridae) | Группа ранних плотоядных ящеротазовых, возможно, ответвившаяся от общего ствола ящеротазовых до того как произошло разделение на теропод и завроподоморф. Иногда классифицируются как тероподы, что в настоящее время оспаривается[58]. |  |
Тероподы (Theropoda)
| Исключительно бипедальные («двуногие») животные с большим разнообразием диет, в том числе птицы. Исходно являлись хищниками, что потребовало адаптации к быстрому бегу и наличия развитого относительно других динозавров интеллекта[c][59]. | .png)
| ||
| † Завроподоморфы (Sauropodomorpha) | Первоначально небольшие бипедальные хищники; продвинутые представители — завроподы (Sauropoda) — преимущественно крупные квадропедальные («четвероногие») растительноядные с длинными шеями и хвостами и колоннообразными ногами | 
| ||
| † Птицетазовые (Ornithischia) | 
| † Гетеродонтозавриды (Heterodontosauridae) |  | |
| † Тиреофоры (Thyreophora) | Включает следующие отчётливо диагностируемые группы:
|  | ||
| † Neornithischia | Включает следующие отчётливо диагностируемые группы:
| 
|
.png)

.png)

| Традиционная топология | «Орнитосцелидная» гипотеза | Гипотеза Бэккера |
|---|---|---|

Эволюционная история
Предки и ранняя эволюция

В настоящее время невозможно определить точный момент, когда появились первые динозавры. Считается, что они отклонились от своих предков, более ранних архозавров, спустя примерно 20 миллионов лет после массового пермского вымирания, которое, по оценкам, уничтожило приблизительно 96% всех морских видов и 70% наземных позвоночных[60][61] Важнейшие обеспечившие успех группе признаки ранних динозавров, отличающие их от большинства других архозавриформ того времени, были связаны с положением тела и характером локомоции. Задние конечности становятся практически вертикальными, а их длина по сравнению с более короткими передними конечностями позволяет делать выводы о передвижении на двух конечностях (бипедальности)[62][63]

По результатам радиоизотопного датирования отложений скальной формации Исчигуаласто[англ.], где были обнаружены ископаемые остатки раннего рода динозавров эораптора (Eoraptor), вероятно относящегося к завроподоморфам[64][65][66], их возраст составляет примерно 231,4 млн лет[67]. Возможно, эораптор напоминал ближайшего общего предка всех динозавров; если это действительно так, то основываясь на его чертах, можно говорить, что первые динозавры были небольшими бипедальными хищниками[68][69][70]. Открытия примитивных, подобных динозаврам орнитодир, таких как Lagosuchus и Lagerpeton[71], из карнийских отложений Аргентины возрастом примерно в 233 млн лет, поддерживают эти предположения; изучение окаменелостей данных рептилий действительно говорит об их малых размерах и вероятном передвижении на двух конечностях.
В 2018 году команда палеонтологов под руководством М. С. Лангера определила, что окаменелости ставрикозавра из отложений формации Санта-Мария[англ.] датируются возрастом в 233,23 млн лет назад, что делает его геологически старше в сравнении с эораптором[72]. Динозавры, возможно, появились еще 243 млн лет назад, о чём свидетельствуют датируемые этим возрастом окаменелости ньясазавра, которые, однако слишком фрагментарны, чтобы уверенно сказать, является ли это животное динозавром или их близким родственником[73]. Ископаемые остатки птицетазовых традиционно считаются плохо задокументированными в триасовых отложениях, но ряд недавних исследований восстановил силезаврид в качестве грады птицетазовых; если силезавриды действительно являются динозаврами, то это древнейшие известные науке представители данной группы[74][75][76].

На самых ранних этапах эволюции динозавров наземная среда обитания была заселена различными группами архозавроморф и терапсид, такими как этозавры, цинодонты, дицинодонты, орнитозухиды, ринхозавры и другие. В конце триаса динозавры выдержали долгую конкуренцию с данными животными, в том числе с во многом более успешными псевдозухиями, прежде чем достигли расцвета. По всей видимости, господствующее положение динозавров было получено не столько превосходством над другими животными, сколько случайностью[77]. Большинство конкурентов ранних динозавров вымерло в триасе в ходе Карнийского плювиального события около 234—232 млн лет назад[78] и триасово-юрского вымирания примерно 201 млн лет назад. Несколько линий ранних динозавров диверсифицировалось в карнийском и норийском веках триаса, предположительно занимая ниши вымерших групп[57][61].
Хронология и палеобиогеография
Послетриасовая эволюция динозавров во многом зависела от изменений во флоре и расположения континентов. В позднем триасе и ранней юре континенты были соединены в единый суперконтинент Пангею, и поэтому существовала всемирная динозавровая фауна, в основном состоявшая из плотоядных целофизоид и ранних завроподоморф[79], постепенно переходивших от хищничества к растительноядности[80]. Голосеменные растения (особенно хвойные), представлявшие потенциальный источник пищи для динозавров, испытывали эволюционную радиацию в позднем триасе[81]. Хотя позднекарнийские завроподоморфы, такие как эораптор и Buriolestes, были хищниками, уже в начале норийского века они начинали переходить на растительноядность и приобретать связанные с ней признаки, такие как длинные шеи[82]. Несмотря на то, что эти растительноядные виды обладали простыми похожими на игуаньи зубами, подходившими прежде всего для пережёвывания мягких листьев, они стали доминирующими растительноядными позднего триаса[81].

Общая однородность динозавровой фауны сохранялась в среднеюрскую и позднеюрскую эпохи, на протяжении которых динозавровая фауна была в основном представлена хищными цератозаврами, мегалозавроидами и карнозаврами и растительноядными стегозаврами и завроподами. Например, такой состав ископаемой фауны наблюдается в отложениях формации Моррисон[англ.] в США и формации Тендагуру[англ.] в Танзании. Динозавры из юрских отложений Китая обладают некоторыми различиями в сравнении с большей частью динозавровой фауны того времени: например, в них обнаружены ископаемые остатки специализированных теропод рода Sinraptor и выделяющихся своими чрезвычайно длинными шеями завропод-мамэньсизаврид[79]. Анкилозавры и орнитоподы становились всё более распространёнными, а базальные завроподоморфы вымерли. Хвойные, папоротниковидные и плауновидные были наиболее распространёнными растениями. Завроподы, как и более ранние завроподоморфы, оставались малоспособными к обработке съеденной пищи в ротовой полости, в то время как птицетазовые развили различные способы пережёвывания, включая подвижные челюсти для обработки пищи и возможные щекоподобные органы для удержания пищи в ротовой полости[81]. Ещё одним заметным эволюционным событием юрского периода стало появление авиал (птиц в широком смысле), произошедших от более ранних целурозавров группы манирапторов[27].
К раннемеловой эпохе благодаря продолжавшемуся распаду Пангеи динозавры стали сильно диверсифицироваться по суше. В самом начале эпохи анкилозавры, игуанодонты и брахиозавриды распространились по Европе, Северной Америке и Северной Африке. Позже представители африканской динозавровой фауны были дополнены или замещены крупными тероподами спинозавридами и кархародонтозавридами, а также завроподами групп реббахизаврид и титанозавров, кроме того известными из отложений соединённой в то время с Африкой Южной Америки. В Азии целурозавры группы манирапторов, такие как дромеозавриды, троодонтиды и овирапторозавры стали типичными представителями теропод, а растительноядные анкилозавриды и ранние цератопсы, такие как пситтакозавр, приобрели роль немаловажных компонентов местных фаунистических комплексов. Между тем Австралия была домом для базальных анкилозавров и орнитопод, а также игуанодонтов[79]. Стегозавры, вероятно, вымерли в какой-то момент в конце раннемеловой эпохи или в начале позднемеловой. Одним из основных изменений для меловой биоты явилось возникновение цветковых растений в раннем мелу и их дальнейшая эволюция, в ходе которой их роль будет усилена в верхнем мелу. Некоторые группы растительноядных динозавров развили более сложноустроенный способ обработки пищи внутри ротовой полости в виде особо типа зубного ряда, называемого зубной батареей, в котором отдельные зубы плотно прилегали друг к другу, формируя непрерывную шлифовальную поверхность. Этот тип независимо появился у цератопсов, игуанодонтов, у которых он был доведён до совершенства[81], и некоторых завропод, среди которых ярким примером является реббахизаврид нигерзавр[83].

В позднем мелу существовало три основных типа динозавровой фауны. На северных континентах, то есть в Северной Америке и Азии, среди теропод преобладали тираннозавриды и различные группы более мелких в сравнении с первыми манирапторов, а местная растительноядная мегафауна была представлена птицетазовыми динозаврами, преимущественно гадрозавридами, цератопсами, анкилозавридами и пахицефалозаврами. На южных континентах, ранее входивших в к тому времени уже разделившуюся Гондвану, тероподы абелизавриды являлись типичными хищными динозаврами, а завроподы титанозавры — типичными растительноядными. В Европе преобладали дромеозавриды, игуанодонты Rhabdodontidae, анкилозавры нодозавриды и завроподы титанозавры[79]. Цветковые растения претерпевали эволюционное излучение[81], в ходе которого к концу мелового периода появились первые травы[84]. Измельчавшие своими клювами растительность гадрозавриды и «состригавшие» её цератопсы стали очень разнообразными в Северной Америке и Азии. Тероподы также диверсифицировались, в том числе увеличилось разнообразие растительноядных и всеядных видов, в особенности теризинозавров и орнитомимозавров[81].
Мел-палеогеновое вымирание, произошедшее примерно 66 миллионов лет назад и ознаменовавшее конец мелового периода и всей мезозойской эры, привело к исчезновению всех групп динозавров, за исключением возникших в позднем мелу веерохвостых птиц (кроновая группа птиц). Некоторые другие рептилии, такие как крокодилы, себекозухии, черепахи, лепидозавры и хористодеры, также пережили это вымирание[85].
Выжившие эволюционные ветви птиц, в том числе бескилевые, гусеобразные и курообразные (Galloanseres), а также ржанкообразные, быстро диверсифицировались в начале палеогенового периода, заняв экологические ниши, оставшиеся свободными в результате исчезновения групп мезозойских динозавров, таких как древесные энанциорнисовые птицы, водные гесперорнисообразные и даже более крупные наземные тероподы, которых в некоторой степени смогли заместить некоторые бескилевые, гасторнисы, дроморнитиды, фороракосовые, Bathornithidae[англ.], Eogruidae[англ.] и Geranoididae[англ.]. Хотя часто утверждается, что млекопитающие превзошли птиц за доминирование в основной массе наземных ниш, многие из этих групп сосуществовали с богатой фауной млекопитающих на протяжении большей части кайнозойской эры[86]. Фороракосовые и Bathornithidae занимали ниши крупных хищников наряду с плотоядными млекопитающими[87][88], а бескилевые всё ещё довольно успешны в качестве растительноядных животных среднего размера; Eogruidae также существовали от эоцена до плиоцена и вымерли только в плиоцене, после более чем 20 млн лет сосуществования со многими группами млекопитающих[89].
Анатомия и физиология
Отличительные анатомические признаки

В настоящее время открытия большого видового разнообразия привели к тому, что представить общепринятый отличительных черт динозавров стало затруднительно; почти всех обнаруженных на сегодняшний день динозавров разделяют определённые изменения по отношению к наследственному от более ранних архозавроморф скелету. Хотя некоторые более поздние группы динозавров показали признаки дальнейшего изменения общих черт, они считаются типичными динозаврами; такие общие черты между различными таксономическими группами называются синапоморфиями[90].
Детальная оценка филогенетических взаимоотношений архозавров Стерлинга Несбитта[англ.] подтвердила или обнаружила следующие двенадцать однозначных синапоморфий динозавров, некоторые из которых были известны ранее[91]:
- Перед надвисочным отверстием присутствует надвисочная ямка;
- Позади первых двух шейных позвонков, атланта и эпистрофея, присутствуют костные выступы эпипофизы;
- Лучевая кость короче плечевой кости на 80 %;
- Четвёртый вертел на бедренной кости представляет собой острый выступ;
- Четвёртый вертел асимметричный, с дистальным нижним краем, образующим более крутой угол к диафизу;
- На таранной и пяточной костях проксимальная суставная фасетка (верхняя соединительная поверхность) малоберцовой кости занимает менее 30% поперечной ширины элемента;
- Внезатылочные кости отсутствуют вдоль средней линии на нижней части эндокринной полости;
- В области таза проксимальные суставные поверхности седалищной кости с подвздошной и лобковой костями отделены большой вогнутой поверхностью (на верхней стороне седалищной кости часть открытого тазобедренного сустава расположена между местом соединения с лобковой и подвздошной костями);
- Кнемиальный гребень на большеберцовой кости дугообразно изгибается антеролатерально (спереди и с внешней стороны);
- Отчётливый проксимодистально ориентированный (вертикальный) гребень присутствует на задней грани дистального конца большеберцовой кости;
- Вогнутая суставная поверхность места соприкосновения малоберцовой и пяточной костей (верхняя поверхность пяточной кости, где она соприкасается с малоберцовой костью, имеет полый контур).
Несбитт представил ряд дальнейших потенциальных синапоморфий и отбросил ряд синапоморфий, предложенных ранее. Некоторые из них также присутствуют у силезаврид, которых Несбитт восстановил в качестве сестринской группы динозавров, в том числе большой передний вертел, плюсневые кости II и IV почти равной длины, уменьшенная степень соединения между седалищной и лобковой костями, наличие кнемиального гребня на голени и восходящего отростка на астрагале, а также многие другие[30]. Тем не менее, в некоторых недавних филогенетических анализах силезавриды были восстановлены как града базальных птицетазовых динозавров[74][75][76].

Множество других скелетных особенностей характерны для всех динозавров. Однако, поскольку они присущи и другим группам архозавров, либо отсутствовали у некоторых ранних динозавров, эти особенности не считаются синапоморфиями. Например, как и большинство диапсид, динозавры имеют две пары подвисочных отверстий и, как представители архозавров, обладают дополнительными отверстиями в передней части черепа и нижней челюсти[92]. Кроме того, известно, что некоторые особенности, которые раньше считались синапоморфиями, возникли до появления динозавров или отсутствовали у самых ранних динозавров и независимо развились у различных групп динозавров. К ним относятся удлинённая лопатка, крестец, состоящий из трёх или более сросшихся позвонков (три встречаются у некоторых других архозавров, но у герреразавра их только два)[30] и перфорированная вертлужная впадина или тазобедренный сустав с отверстием в её центре внутренней поверхности (к примеру, закрыто у сатурналии)[93][94]. Другая трудность определения чётко выраженных особенностей динозавров состоит в том, что ранние динозавры и другие архозавры позднего триаса часто плохо известны и во многих отношениях были похожи; эти животные иногда неправильно идентифицировались в литературе[95].
Размер


Имеющиеся данные свидетельствуют о том, что средний размер динозавров весьма варьировался на протяжении мезозойской эры[69]. По результатам исследования Джеймса Фарлоу 1993 года, плотоядные динозавры-тероподы, которые занимали большинство ниш наземных хищников в течение мезозоя, при сортировке по категориям порядка величины на основе предполагаемого веса чаще всего попадают в категорию весящих от 100 до 1000 кг, тогда как крупнейшие современные хищные хищные млекопитающие входят в категорию весящих от 10 до 100 кг[96]. Мода массы тела нептичьих динозавров, согласно оценке Яна Печкиса 1995 года, составляла 1—10 т[97]. Это резко контрастирует со средним размером кайнозойских млекопитающих, оцененным Национальным музеем естественной истории, Вашингтон, примерно в 2—5 кг[98].
К числу динозавров-завропод относились самые крупные и тяжёлые когда-либо существовавшие наземные животные, некоторые из которых, вероятно, по длине превышали самых больших китов, хотя и уступали им по массе[99][100][101]. Своих размеров завроподы, по всей видимости, достигли за счёт сочетания быстрого роста, длинной шеи, эффективных лёгких и системы воздушных мешков, предназначенной для облегчения массы и охлаждения тела[99]. Предполагается несколько возможных преимуществ большого размера для завропод, в том числе защита от хищников, сокращение потребления энергии и большая продолжительность жизни, но, возможно, самым важным из них была роль в питании. Крупные животные более эффективно переваривают пищу, чем мелкие, потому что пища проводит больше времени в их пищеварительной системе. Это также позволяет им питаться пищей с более низкой питательной ценностью, чем это возможно для более мелких животных. Ископаемые остатки завропод в основном встречаются в геологических формациях, которые интерпретируются как сухие или сезонно засушливые, и способность питаться большими количествами низкокалорийной пищи была бы выгодна в таких условиях[102].
Рекордные размеры
.jpg)
По всей видимости, никогда не представится возможным достоверно установить, какой вид динозавров был действительно самым большим по длине, высоте или массе. Кандидаты на перечисленные статусы представлены сравнительно фрагментарным материалом, причём оценки их размера, которые могут весьма варьировать, дают возможность определить размер только конкретной особи, а также не позволяют исключать вероятность открытия ещё большего динозавра в будущем[103][104][105].
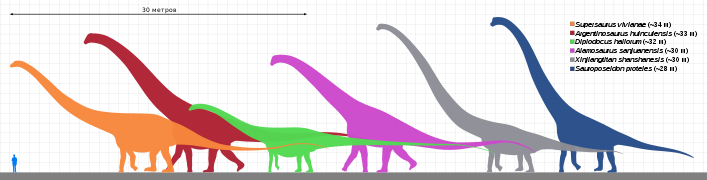
Жираффатитан (Giraffatitan) является самым высоким и самым тяжёлым динозавром, известным по полному скелету[106][107]; согласно оценке Х. Маццетты 2004 года, взрослое животное могло достигать 21,8 м при массе в 39,5 т[108]. Относительно полным материалом представлены диплодок Карнеги (Diplodocus carnegii) и Diplodocus hallorum, оцененные Г. С. Полом[англ.] в 2016 году как достигавшие 24 м при массе в 12 т и и 29 м при массе в 23 т соответственно[109]. По оценке Пола 2019 года, известные по более фрагментарному материалу аргентинозавр (Argentinosaurus) и патаготитан (Patagotitan) могли достигать примерно 35 м при массе в 70 т и 31 м при массе в 52 т соответственно[110]. Кроме того, в число одних из крупнейших динозавров входят суперзавр (Supersaurus)[111], а также, возможно, барозавр (Barosaurus)[112][113] и известный по утерянной дуге позвонка Maraapunisaurus fragillimus[114].
Одним из самых больших плотоядных животных, живших на Земле, был динозавр-теропод спинозавр (Spinosaurus), реконструкция скелета которого, расположенная в Национальном географическом музее (National Geographic Museum), достигает 15 м в длину[115][116]. К крупнейшим тероподам также относились тираннозавр (Tyrannosaurus), гиганотозавр (Giganotosaurus), кархародонтозавр (Carcharodontosaurus) и мапузавр (Mapusaurus), достигавшие примерно 12—13 м в длину[117]; так, в 2020 году Скотт Персонс и коллеги оценили длину выделяющегося своими размерами образца тираннозавра RSM P2523.8 («Скотти)», полного примерно на 65%, в 13 м, а его массу — в 8,8 т[118][119].
Самый маленький из известных науке представителей клады динозавров — колибри-пчёлка (Mellisuga helenae), для которой обычны длина в 5 см и масса в 2 г[120][121]. Самым маленьким известным мезозойским динозавром является энанциорнисовая птица Cratoavis?!, достигавшая 6,6 см в длину при массе приблизительно в 4 г (примерно в 2,1 млн раза меньше массы тираннозавра), а самым маленьким научно описанным нептичьим тероподом — эпидексиптерикс (Epidexipteryx), достигавший 25 см при массе примерно в 220 г[121].
Крупнейшим известным птицетазовым является гадрозаврид шаньдунозавр (Shantungosaurus), который, по оценке Пола (2016), достигал 15 м при массе в 13 т[122].
 Некоторые гигантские завроподы
Некоторые гигантские завроподы Некоторые гигантские тероподы
Некоторые гигантские тероподы Некоторые гигантские орнитоподы
Некоторые гигантские орнитоподы Самые маленькие нептичьи тероподы
Самые маленькие нептичьи тероподы Колибри-пчёлка является самым маленьким известным науке представителем клады динозавров
Колибри-пчёлка является самым маленьким известным науке представителем клады динозавров




.svg)
Метаболизм
Поскольку как современные крокодилы, так и птицы, имеют четырёхкамерные сердца (хотя и модифицированные у крокодилов), вполне вероятно, что эта черта присуща всем архозаврам, включая всех динозавров[123].

Все современные птицы обладают высоким метаболизмом и являются эндотермными («теплокровными»); с 1960-х годов ведутся активные дебаты относительно того, насколько сильно эта черта распространена в родословной динозавров. Различные авторы считали нептичьих динозавров экзотермными («холоднокровными»), мезотермными[англ.] (обладавшими промежуточным типом метаболизма) или эндотермными, причём первый вариант хотя и принимался за факт в прошлом, в современной литературе больше не воспринимается всерьёз[124].
Среди исследователей формируется консенсус о том, что, хотя разные линии динозавров имели разный метаболизм, у некоторых или многих из них уровень метаболизма был выше, чем у других нептичьих рептилий, но ниже, чем у современных птиц и млекопитающих (мезотермия)[125][126][127]. Из современных животных мезотермами являются ехидновые, кожистые черепахи, тунцы и акулы[127]. Данные, полученные благодаря изучению крокодилов и их вымерших родственников, предполагают, что повышенный метаболизм мог развиться у самых ранних архозавров, являвшихся общими предками динозавров и крокодилов[128][129].

Исторически нептичьи динозавры рассматривались в качестве эктотермов. Считалось, что они были относительно медленными, вялыми животными, несмотря на то, что многие современные рептилии довольно быстрые и ловкие, хотя и полагаются на внешние источники тепла для регулирования температуры своего тела. Мнение об эктотермии динозавров оставалось преобладающей точкой зрения до тех пор, пока Роберт Бэккер, один из первых сторонников эндотермии динозавров, не опубликовал влиятельную статью на эту тему в 1968 году. Бэккер использовал анатомические и экологические данные, чтобы доказать, что завроподы, которые на тот момент изображались как медлительные водные животные с волочащимися по земле хвостами, были эндотермами, ведшими активный наземный образ жизни. В 1972 году Бэккер дополнил свои аргументы, основываясь на предполагаемых потребностях в энергии и принципах строения системы «хищник — жертва». Эти работы стали одними из основополагающих для современной палеонтологии динозавров и ознаменовали начало Ренессанса динозавров[англ.] (см. § Ренессанс динозавров)[130][131][132][133].
Значительный вклад в современное понимание физиологии динозавров внесла палеогистология, изучение микроскопической структуры тканей вымерших организмов[134][135]. Начиная с 1960-х годов и далее Арман де Риклес[англ.] предполагал, что наличие у динозавров фиброламеллярной кости — костной ткани с неправильной, волокнистой текстурой, заполненной кровеносными сосудами — свидетельствует о стабильно быстром росте и, следовательно, эндотермии. Фиброламеллярная кость была распространена как среди динозавров, так и среди птерозавров[136][137], хотя не все из них обладали ей[138][139]. Деятельность де Риккла показала актуальность палеогистологического анализа для решения серьёзных эволюционных вопросов, что повлекло за собой публикации множество новых работ по реконструкции кривых роста и моделированию эволюции темпов роста в различных линиях динозавров, вместе взятые результаты которых предполагают, что динозавры росли быстрее, чем современные нептичьи рептилии[135].

К другим свидетельствам, говорящим об эндотермии нептичьих динозавров, относятся наличие перьев и других типов покровов на теле у многих их групп, более постоянные соотношения изотопа кислорода-18 в костной ткани по сравнению с эктотермами, особенно по мере измерения географической широты и, следовательно, температуры воздуха, что предполагает стабильные внутренние температуры (хотя эти соотношения могут изменяться при фоссилизации[140])[141][142], и открытие полярных динозавров, живших на территории нынешних Австралии, Антарктиды и Аляски, когда в этих местах был холодный умеренный климат[143][144][145][146].
Некоторые оценки интенсивности метаболизма динозавров показывают, что двуногие динозавры массой 20 кг и более в любом случае были теплокровными[147]. Однако, спорным вопросом является способ поддержания температуры тела. Свидетельство того, что крупные динозавры были инерциальными эндотермами, получили в 2006 году Д. Гиллули, Э. Аллен и Э. Чарнов. Исследовав рост костей крупных динозавров, они выяснили, что в начале жизни температура тела крупных динозавров составляла +25 °C, а по мере достижения массы в несколько сотен килограммов она значительно вырастала и достигала у самых крупных экземпляров +35 °C. В то же время у мелких динозавров с ростом температура тела не увеличивалась. Был сделан вывод, что по мере увеличения размеров возникает способность удерживать тепло.
Покровная система

Хотя нептичьи динозавры и древние птицы в основном представлены скелетным материалом, известны также отпечатки их чешуи, нитевидных структур и перьев, свидетельствующие о высоком разнообразии покровов среди динозавров. Существует ряд образцов динозавров с отпечатками чешуи высокой сохранности. Для многих нептичьих динозавров была характерна чешуйчатая кожа, состоявшая из плотно располагавшихся друг к другу мелких чешуек, среди которых иногда встречались чешуйки пирамидальной формы, обладавшие бо́льшим размером[149]. Тем не менее, не существовало какого-либо определённого типа динозавровой чешуи: даже у близкородственных видов часто наблюдаются значительные различия в её структуре[150].


Известны многочисленные отпечатки чешуи гадрозаврид, благодаря чему стало возможным оценить различия в покрове разных видов, который, в некоторых случаях, удалось практически полностью реконструировать[151].
В ряде случаев по ископаемым остаткам, содержащим пигмент, удаётся установить цвета окраса динозавров[152].
Перья и протоперья

Перья, одна из самых узнаваемых черт современных птиц, также присутствовали у ряда нептичьих динозавров. Прямые свидетельства наличия перьев или перьеподобных структур были обнаружены на ископаемых остатках многих теропод группы целурозавров, причём отпечатки истинных, лопастных перьев, похожих на маховые перья современных птиц, были найдены только у манирапторов. Отпечатки перьеподобных структур обнаружены у птицетазовых, а именно у тяньюйлуна (Tianyulong)[154], кулиндадромея (Kulindadromeus)[155] и пситтакозавра (Psittacosaurus)[156], а также птерозавров (пикнофибры); если они гомологичны перьям, что является дискуссионным вопросом[157], то это означает, что примитивные перья возникли ещё до динозавров и имелись уже у ранних авеметатарзалий[148].
Ископаемые остатки археоптерикса (Archaeopteryx) с отпечатками перьев, обнаруженные в зольнхофенском известняке, Германия, стали первой окаменелостью, наглядно показавшей связь между динозаврами и птицами. Открытие этого животного в 1859 году, всего через два года после публикации основополагающего труда Чарлза Дарвина «Происхождение видов», вызвало зарождавшиеся дебаты между сторонниками эволюционной биологии и креационизма. Начиная с 1990-х годов были описаны отпечатки перьев и перьеподобных структур нептичьих динозавров из формаций Исянь[англ.], Цзюфотан[англ.] и Тяоцзишань[англ.] (Ляонин, Китай)[158], Крато[англ.] (северо-восток Бразилии)[159], Дайносор-Парк[англ.] (Альберта, Канада)[160][161] и укурейской свиты[англ.] (Забайкальский край, Россия)[155]; в бирманском янтаре (Мьянма) был найден хвост целурозавра с сохранившимися перьями, принадлежавший нептичьему представителю клады, либо древней птице[162].

По всей видимости, на ранних стадиях эволюции перьев у теропод основными факторами отбора были потребность в теплоизоляции[148] и полезность их использования для демонстрации. Сюй Син и Ю Го (2009) выделяют следующие стадии эволюции пера[163]:
- Одиночное волокно;
- Множество волокон, соединённых у их основания;
- Множество волокон, соединённых у основания центральным волокном;
- Множество волокон, прикреплённых к центральному волокну;
- Множество волокон, поднимающихся из края мембранной структуры;
- Контурное перо[англ.] с лопастью из бородок и центральной осью;
- Контурное перо с ассиметричным стержнем;
- Недифференцированная лопасть со стержнем по центру.
Подтверждаемое отпечатками присутствие перьев в виде пухообразных ворсинок у базальных тираннозавроид ютираннуса (Yutyrannus) и дилуна (Dilong), но наличие чешуи и отсутствие прямых доказательств существования перьевого покрова у тираннозаврид свидетельствует о том, что тираннозавриды вторично утратили перья в связи с климатическими условиями и, возможно, своим гигантизмом[164]. Палеохудожник Марк Уиттон[англ.] предположил, что вторичная утрата перьев могла иметь место и среди других крупных целурозавров, таких как теризинозавриды и дейнохейриды, теплоэнергетика которых, по его мнению, не требовала наличия густого перьевого покрова[165].
Нервная система
В общих чертах размер и форму мозга можно реконструировать на основе окружающих его костей. В 1896 году Чарльз Марш рассчитал отношения между массой мозга и массой тела семи видов динозавров, показав, что пропорции мозга динозавров по отношению к предполагаемым размерам тела были меньше, чем у современных крокодилов, причём мозг стегозавра был пропорционально меньше, чем у любого современного наземного позвоночного. Это способствовало к популяризации широко распространённого общественного мнения о динозаврах как о медлительных и необычайно глупых животных. Гарри Джерисон в 1973 году показал, что пропорционально меньшие размеры мозга обычно сопровождают большие размеры тела, и что размер мозга у динозавров был не меньше, чем у современных рептилий[166]. Более поздние исследования показали, что относительный размер мозга прогрессивно увеличивался в ходе эволюции теропод. Самый высокий интеллект, сравнимый с интеллектом современных птиц, был рассчитан по черепной коробке троодонтида, которого на тот момент относили к роду Troodon[167] (теперь этот род признаётся nomen dubium, а весь материал, кроме голотипного зуба, распределяется между родами Stenonychosaurus и Latenivenatrix[168][169]).
Дыхательная система

Дыхательная система ящеротазовых, в том числе птиц, характеризуется наличием обширной системы воздушных мешков, то есть воздухоносных полостей во многих костях скелета, делающих их полыми[170]. Воздушные мешки могли появиться у самых ранних ящеротазовых[171]. Они обеспечивали динозавров бо́льшим количеством кислорода по сравнению с млекопитающими аналогичного размера, а также увеличивали уровень дыхательного объёма[англ.] в состоянии покоя и требовали более низкой частоты дыхания, что позволяло поддерживать более высокий уровень активности[172].
Выделительная система
Подобно другим наземным позвоночным, нептичьи динозавры, также как и большинство птиц, являлись урикотеликами, т. е. конечным продуктом их азотистого обмена была мочевая кислота. Выделение мочевой кислоты, а не более токсичного аммиака, позволяет соблюдать гидробаланс, т. к. выведение из организма мочевой кислоты требует минимальных затрат воды (также возможно в твёрдом виде)[173]. У видов птиц мочевая кислота выделяется вместе с фекалиями в виде полутвёрдых отходов[174][175]. Однако, по крайней мере, некоторые современные птицы (например, колибри), являются факультативными аммонотеликами, выделяя большую часть продуктов азотистого обмена в виде аммиака[176]. Продукты азотиского обмена, а также конечные продукты пищеварения, выходят из клоаки[177][178]. Кроме того, многие виды птиц отрыгивают погадки[179]; известны сохранившиеся в ископаемом состоянии погадки позднеюрского манираптора анхиорниса (Anchiornis)[180].
Изучение ископаемых экскрементов, называемых копролитами, позволяет делать выводы о рационе динозавров и их пищеварительных процессах[181][182].
Поведение
Общепринято, что некоторые поведенческие особенности, которые распространены среди птиц, а также у их ближайших современных родственников — крокодилов — были характерны и для нептичьих динозавров[183]. Выводы о поведении динозавров делаются на основании поз, в которых сохранились ископаемые остатки, данных об их среде обитания, путём сравнения их поведения с поведением современных животных, занимающих сходные экологические ниши, и при помощи компьютерного моделирования биомеханики[184].
Рацион и пищевое поведение
.jpg)
Как и у современных птиц, рацион нептичьих динозавров был весьма сложным и неоднородным. Общие представления о нём палеонтологи получают благодаря изучению морфологии зубов и челюстей динозавров, состава их копролитов, содержимого желудка, когда такое сохраняется, а также путём сопоставления следов зубов на костях жертвы с самими зубами. При помощи микроскопов прослеживаются следы износа на зубах (микроизнос), по которым можно определить, как изучаемый динозавр двигал челюстями и какие виды пищи ел[186].
Первые динозавры, скорее всего, были хищниками, но с процессом диверсификации динозавров разные группы специализировались на различных типах питания, благодаря чему они смогли занять многочисленные наземные экологические ниши. В частности, многие крупные тероподы стали гиперплотоядными, продвинутые завроподоморфы и птицетазовые — листоядными[англ.], также некоторые таксоны заняли ниши ихтиофагов (рыбоядных), всеядных и плодоядных[187].
У теропод, как современных, так и вымерших, наблюдается высокое разнообразие диет. Хотя ранние их представители были хищниками, уже на ранних этапах своей эволюции одна из подгрупп теропод, целурозавры, стала очень пластичной в отношении рациона[188][189][190]. Согласно исследованию Л. Занно[англ.] и П. Маковицкого 2011 года, многие целурозавры, в том числе теризинозавры, орнитомимозавры, овирапторозавры, альваресзавриды, ранние авиалы (птицы) и троодонтид джиньфенгоптерикс (Jinfengopteryx), занимали ниши растительноядных и всеядных, тогда как тираннозавриды, компсогнатиды, дромеозавриды и большинство троодонтид — ниши хищников[188]. Следы укусов позволяют сделать вывод о том, что некоторые тероподы, включая майюнгазавра[191], тираннозавра[192] и аллозавра, практиковали каннибализм[193], что наблюдается также и у некоторых современных птиц (хотя клюв затрудняет совершение каннибалистического акта).

Птицетазовые динозавры питались прежде всего различными типами растительности. Те из них, у которых были широкие, почти квадратные морды и нижние челюсти, могли срезать за раз много растительности и, соответственно, не были сильно разборчивы в пище (как белые носороги и бегемоты). Узкими, острыми мордами и челюстями обладали стегозавры и многие орнитоподы; такой тип строения морды свидетельствует об их специализации на каких-либо конкретных типах пищи, таких как молодые побеги, почки или плоды.

Социальность

Современные птицы демонстрируют высокую степень социальности и часто живут в стаях. Первым потенциальным свидетельством стадности некоторых нептичьих динозавров стало открытие в 1878 году 31 скелета игуанодона (Iguanodon) в Берниссаре, Бельгия. Как было предположено, животные утонули после того, как они упали в глубокую затопленную карстовую воронку[194]. Впоследствии были обнаружены и другие массовые захоронения остатков. Это, наряду с множеством тропинок, предполагает, что стадное поведение было обычным явлением для мезозойских динозавров. Следы сотен или даже тысяч растительноядных животных указывают на то, что орнитоподы гадрозавриды, возможно, передвигались большими стадами, как бизоны или антилопы спрингбоки.
Следы завропод, обнаруженные в Оксфордшире, Англия, документально подтверждают, что оставившие их животные путешествовали группами, состоявшими из нескольких разных видов[195], хотя при этом нет никаких доказательств существования какой-либо особой структуры стада. Объединение в стада могло быть связано с вытекающей отсюда большей безопасностью, в том числе для молодняка, и с возможностью осуществления массовых миграций. Есть свидетельства того, что многие клады динозавров, которые росли медленно, такие как различные тероподы, завроподы, анкилозавры, орнитоподы и цератопсы, образовывали группы из незрелых особей. Одним из примеров являются обнаруженные во Внутренней Монголии ископаемые остатки более 20 особей Sinornithomimus, погибших в возрасте от одного года до семи лет. Эта находка интерпретируется как социальная группа, погибшая, застряв в грязи.
Интерпретация растительноядных динозавров как стадных животных также распространилась на изображение плотоядных нептичьих теропод в качестве стайных охотников[англ.], совместно нападавших на крупную добычу[196][197]. Однако такой вид охоты редок среди современных птиц, крокодилов и других завропсид, а тафономические данные, предполагающие стайную охоту, как у млекопитающих, у таких теропод как дейноних и аллозавр (Allosaurus), можно также истолковать как результаты битв на смерть за тушу, встречающихся и среди многих современных плотоядных завропсид[198].
Способы коммуникации

Среди нептичьих динозавров наблюдается большое разнообразие «причудливых структур» (англ. bizarre structures), таких как воротники и купола на головах цератопсов и пахицефалозавров соответственно, рога, гребни и паруса; вероятно, они предназначались для внутривидовой (возможно, межполовой) коммуникации[199][200]. По мнению В. С. Персонса и Ф. Дж. Карри (2019), на ранних своих этапах эволюция перьев протекала под непосредственным влиянием полового отбора[201].
Звуковые сигналы имеют крайне высокое значение в жизни современных птиц, но их роль в коммуникации нептичьих динозавров более спорна. В 2008 году палеонтолог Фил Сентер предположил, что нептичьи динозавры полагались в основном на визуальные образы и, возможно, неголосовые акустические звуки, такие как шипение, скрежет челюстями или, в случае с манирапторами, на хлопки крыльями. Сентер заявил, что они вряд ли были способны к вокализации, поскольку их ближайшие родственники, крокодилы и птицы, используют разные средства вокализации: первые издают звуки гортанью, а вторые — через уникальный орган сиринкс. По мнению Сентера, сиринкс возник независимо от гортани и, соответственно, общий предок птиц и крокодилов был немым[202].
Самые ранние известные минерализированные остатки сиринкса были описаны в 2016 году командой палеонтологов под руководством Джулии Кларк[англ.]; они принадлежали верхнемеловой птице вегавису (Vegavis), жившей примерно 69—66 миллионов лет назад. По мнению Кларк и коллег, сиринкс вряд ли существовал у нептичьих динозавров, но, тем не менее, они всё же были способны к вокализации. Команда предположила, что голосовая система птиц, основанная на сиринксе, эволюционировала из системы, основанной на гортани[203]. Исследование, проведённое в 2016 году, показало, что нептичьи динозавры, вероятно, издавали звуки с закрытым ртом, которые встречаются как у крокодилов, так и у птиц, а также у других завропсид (в частности, могли ворковать). Способность к такой вокализации неоднократно независимо развивалась у архозавров после увеличения размеров тела, что подтверждается их современными представителями[204].
Предполагается, что гребни ламбеозаврин и носовые камеры анкилозаврид исполняли роль резонирующих камер, предназначенных для получения низкочастотных звуков[205][206]; Сентер, однако, отмечает, что наличие резонирующих камер у некоторых динозавров не обязательно свидетельствует об их способности к вокализации, поскольку у современных змей есть аналогичные камеры, которые усиливают их шипение[202].
Территориальность
Гребни и «воротники» некоторых динозавров, таких как маргиноцефалы, ламбеозаврины и некоторые тероподы, скорее всего, были слишком хрупкими для использования в целях активной защиты, и поэтому они, вероятно, использовались для демонстрационных целей, чтобы напугать соперника, либо привлечь внимание партнёра для спаривании, хотя мало что известно о спаривании нептичьих динозавров и их территориальности. Раны от укусов на черепах говорят о том, что тероподы регулярно принимали участие в схватках[207].
Защита
Имеются доказательства того, что некоторые виды небольших птицетазовых динозавров (например, небольшое птицетазовое Oryctodromeus cubicularis) выкапывали себе норы. Данная находка была сделана в 2005 году в штате Монтана. Палеонтологи обнаружили не только их норы, но и сами кости динозавров. Изучение костей показало, что динозавры по какой-то причине умерли в своём убежище, а потом оно оказалось засыпано, возможно, во время наводнения. В 2009 году в Австралии повторно были обнаружены норы динозавров. До открытия нор палеонтологи предполагали, что динозавры спали где-то под корнями деревьев и в других естественных убежищах. Теперь же учёные предполагают, что некоторые виды динозавров выкапывали себе норы как раз для спячки[208].
Размножение

В общих чертах половое поведение мезозойских динозавров может быть установлено путём филогенетического брекетинга — многие особенности поведения, которые характерны для птиц и крокодилов, вероятно, были характеры и для вымерших архозавров. Не вызывает сомнений тот факт, что динозаврами практиковалось внутреннее оплодотворение. Строительство гнёзд и кладка яиц подтверждаются также ископаемыми остатками. Вероятно, большинство динозавров использовало стратегию r-отбора, то есть рождали многочисленных детёнышей, из которых лишь немногие доживали до взрослого возраста; это подтверждается существованием отложениями, где большая часть следов оставлена молодыми особями, а также многочисленными остатками не закончивших свой рост динозавров[209].
Когда самки птиц начинают производить яйца, у них возникает медуллярная кость — особый тип костной ткани в форме губчатой массы, появляющийся между твёрдой наружной костью и костным мозгом задних конечностей. В медуллярной кости накапливается кальций для формирования скорлупы яиц. Таким образом, наличие медуллярной кости позволяет узнать, что животное было половозрелой самкой; это может помочь в определении пола образцов мезозойских динозавров. Подобная медуллярной кости ткань была идентифицирована у мезозойских теропод тираннозавра, аллозавра и конфуциорниса (ранняя птица) и птицетазового тенонтозавра[210], а также наличие такой указывалось для завроподоморфа мусзавра. Если идентификация верна (что является спорным вопросом[211]), то медуллярная кость имелась у всех линий динозавров. Примечательно, что особи с предполагаемым наличием медуллярной кости были подростковыми, а значит их половая зрелость наступала до того, как животные достигали своего полного размера[210].
Гениталии динозавров вряд ли когда-нибудь будут обнаружены в ископаемом состоянии, поскольку на них отсутствовали чешуйки, которые могли бы обеспечить сохранение за счёт пигментации или остаточных солей фосфата кальция. Половые органы мезозойских динозавров находились в клоаке, как у птиц и крокодилов. В 2021 году была подробно описана внешняя часть клоаки пситтакозавра (Psittacosaurus sp.), которая как по форме, так и по строению, оказалась близка к таковой у крокодилов. Клоака пситтакозавра представляла собой продольное отверстие, окружённое морщинистыми тканями. По мнению авторов описания, пигментация клоаки может говорить о том, что, возможно, с помощью неё подавались сигналы особям противоположного пола; аналогичное поведение наблюдается и у современных птиц, как самок, так и самцов, что делает невозможным определение пола образца[212].
Яйца и гнёзда


И крокодилы, и птицы, откладывают амниотические яйца и в той или иной мере заботятся о потомстве; аналогичная ситуация наблюдалась и у мезозойских динозавров, что подтверждается не только филогенетическим брекетингом, но и палеонтологическим материалом. Обнаружены многотысячные окаменелости яиц динозавров, причём в большинстве случаев не представляется возможным определить, какому конкретному виду они принадлежат. В отдельных случаях в яйцах сохраняются остатки эмбрионов[213]. Яйца многих нептичьих динозавров были покрыты твёрдой скорлупой, в значительной мере состоявшей из карбоната кальция, как у птиц и крокодилов. Тем не менее, по результатам исследования М. Норелла[англ.] и коллег (2020), первые динозавры откладывали яйца с мягкой скорлупой, как у черепах, а твёрдая скорлупа как минимум трижды независимо приобреталась в основных группах динозавров (птицетазовые, завроподоморфы и тероподы)[214].
В 2005 году из верхнего мела Китая был описан овирапторозавр с двумя неотложенными яйцами в тазовом канале; исходя из количества яиц, был сделан вывод о том, что у большинства нептичьих теропод, как и крокодилов, имелось два функциональных яйцевода, но, как у птиц, каждый из них производил только одно яйцо за раз (вероятно, самка откладывала по два яйца в день). У современных птиц функциональным является только левый яйцевод[215].
Среди птиц и крокодилов широко распространена забота о потомстве; вероятно, эта особенность поведения имелась уже у первых архозавров и была унаследована их потомками, в том числе динозаврами[183]. В 1979 году Д. Р. Хорнер и Р. Макела описали коммунальное гнездовье майазавр (Maiasaura, буквально — «ящер-хорошая-мать») из верхнего мела Монтаны, продемонстрировав наличие длительной заботы за птенцами после их вылупления по меньшей мере у некоторых орнитопод-гадрозаврид[216]. Следы с острова Скай на северо-западе Шотландии также подтвердили наличие родительского поведения среди орнитопод[217]. Образец овирапторида читипати (Citipati)[218] из верхнего мела Монголии был обнаружен в положении, как у курицы-наседки[219][220], что может указывать на то, что уже тогда манирапторы использовали изолирующий слой перьев для согревания яиц[221]. У эмбриона базального завроподоморфа массоспондила (Massospondylus) отсутствуют зубы, из чего следует, что после вылупления детёнышам этих динозавров требовалась родительская забота[222].
Существует также ряд доказательств выводковости или полувыводковости многих видов динозавров, особенно теропод. Например, не относящиеся к кладе Ornithuromorpha мезозойские птицы имели медленные темпы роста, закапывали яйца, как нынешние большеноги, и были способны летать вскоре после рождения[223][224][225][226]. И тираннозавр, и стенонихозавр, явно были выводковыми, а их детёныши, вероятно, занимали экологические ниши, отличные от таковых у взрослых особей[227]. Также был сделан вывод о выводковости завропод[228].
В 2010 году был описан район колониальных гнездовий неозавропод, представляющий собой доказательство того, что по крайней мере некоторые динозавры-завроподы устраивали паровой обогрев кладок яиц, на постоянной основе откладывая их возле геотермальных источников. Схожее поведение наблюдается у современного полинезийского большенога (Megapodius pritchardii), вместо насиживания яиц закапывающего их в тёплые вулканические пески и почву[229].
 Кладка овираптора
Кладка овираптора Образец высиживавшего яйца читипати. Окаменелость справа представляет собой яйцо в разрезе.
Образец высиживавшего яйца читипати. Окаменелость справа представляет собой яйцо в разрезе. Модель-реконструкция гнезда майазавры
Модель-реконструкция гнезда майазавры Кустарниковый большеног на его закрытом гнезде. Предполагается, что аналогичные гнёзда могли строить некоторые нептичьи динозавры.
Кустарниковый большеног на его закрытом гнезде. Предполагается, что аналогичные гнёзда могли строить некоторые нептичьи динозавры.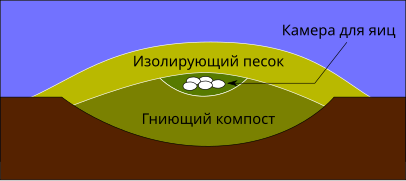 Разрез гнезда большеногов
Разрез гнезда большеногов
.jpg)




Вымирание основных групп
Все нептичьи динозавры и большинство ветвей птиц (авиал)[230] вымерли в результате произошедшего в конце мелового периода массового вымирания, называемого мел-палеогеновым вымиранием. Выше мел-палеогеновой границы, которая датируется возрастом в 66,038 ± 0,025 млн лет[231], окаменелости нептичьих динозавров внезапно исчезают; их отсутствие исторически использовалось для отнесения пород к последующей кайнозойской эре. Причины массового вымирания обсуждаются с 1970-х годов, что привело к развитию идей о том, что важную в роль в нём сыграли импактное событие, заключавшееся в падении крупного метеорита на территорию полуострова Юкатан, и повышенный вулканизм в Деканских траппах, Индия. Тем не менее, особенности механизмов вымирания и степень его воздействия на динозавров являются объектами продолжающихся исследований[232]. Наряду с большинством ветвей динозавров вымерли многие другие группы животных: птерозавры, различные морские рептилии, такие как мозазавры и плезиозавры, несколько ветвей млекопитающих, головоногие моллюски аммониты, принимавшие участие в образовании рифов двустворчатые моллюски рудисты и различные группы морского планктона[233][234]. Всего в ходе мел-палеогенового вымирания на Земле вымерло примерно 47% родов и 76% видов[235]. Относительно крупные размеры большинства нептичьих динозавров и низкое разнообразие небольших их представителей в конце мелового периода, возможно, способствовали их вымиранию[236]; вымирание многих линий птиц могло быть вызвано их зависимостью от лесных местообитаний или отсутствием полезной для выживания в условиях вымирания адаптации к поеданию семян[англ.][237][238].
Разнообразие перед вымиранием
Разнообразие нептичьих динозавров, существовавших во всём мире непосредственно перед мел-палеогеновым вымиранием, оценивается в 628—1078 видов[239]. Остаётся неясным, сокращалось ли оно постепенно вплоть до исчезновения в ходе мел-палеогенового вымирания, или же динозавры процветали до вымирания. Было обнаружено, что горные формации маастрихтского яруса, отложившиеся в самом его конце, перед вымиранием, содержали меньшее разнообразие остатков нептичьих динозавров, чем формации предшествующего кампанского яруса, что привело к преобладанию мнения о постепенном сокращении разнообразия[240][241][242]. Однако в этих сравнениях не учитывалось варьирование степени сохранности остатков в разных отложениях, а также опускалась неравномерная изученность этих отложений. В 1984 году Дейл Рассел[англ.] не обнаружил никаких доказательств сокращения разнообразия[243]. Другой анализ, проведённый Дэвидом Фастовским и его коллегами в 2004 году, даже показал, что разнообразие динозавров постоянно увеличивалось вплоть до мел-палеогенового вымирания[244], но выводы этого анализа были оспорены Робертом Салливаном в 2006 году[245]. С тех пор различные подходы, основанные на статистике и математических моделях, по-разному поддерживали либо версию о внезапном вымирании[232][239][246], либо о постепенном снижении разнообразия[247][248]. Тенденции разнообразия в конце мелового периода, возможно, варьировались между разными группами динозавров: было высказано предположение, что завроподы не испытывали спада разнообразия, в то время как птицетазовые и тероподы приходили в упадок[249][250].
Импактное событие

Гипотеза, впервые представленная научному сообществу в 1980 году Уолтером Альваресом, Луисом Альваресом и их коллегами, объясняет мел-палеогеновое вымирание столкновением с Землёй болида (чрезвычайно яркого метеора). Альварес и его коллеги предположили, что резкое увеличение уровней иридия, зафиксированное во всем мире в горных отложениях на мел-палеогеновой границе, является прямым свидетельством такого столкновения[251]. Кроме того, по всему миру обнаружен шоковый кварц[англ.], свидетельствующий о сильной ударной волне, исходившей от удара[252]. Фактическое место падения оставалось неизвестным, пока на полуострове Юкатан, расположенном на юго-востоке Мексики, не был обнаружен кратер Чикшулуб шириной в 180 км, о чём Алан Хильдебранд[англ.] и коллеги сообщили в опубликованной в 1991 году статье[253]. Теперь бо́льшая часть свидетельств позволяет предполагать, что болид шириной от 5 до 15 км врезался в полуостров Юкатан около 66 млн лет назад, образовав этот кратер[254] и создав «механизм уничтожения», который вызвал вымирание[255][256][257].

В течение нескольких часов после столкновения с метеором должен был произойти ряд глобальных стихийных бедствий, таких как землетрясения[258], цунами[259] и огненный смерч, который, вероятно, убил незащищённых животных и вызвал лесные пожары[260][261]. Однако также должны были возникнуть и долгосрочные последствия для окружающей среды. На протяжении нескольких дней сульфатные аэрозоли, выбрасываемые из горных пород в месте падения, будут способствовать возникновению кислотных дождей и закислению океана[262][263]. Предполагается, что аэрозоли сажи распространялись по всему миру в последующие за столкновением месяцы и годы, охладив поверхность Земли за счёт отражения теплового излучения и значительно замедлив фотосинтез, блокируя солнечный свет, тем самым вызвав импактную зиму[232][264][265] (роль в возникновении импактной зимы приписывалась сульфатным аэрозолям, пока эксперименты не продемонстрировали несостоятельность этого предположения[263]). Прекращение фотосинтеза подразумевало разрушение пищевых сетей, зависящих от лиственных растений, частью которых были и все динозавры, за исключением зерноядных птиц[238].
Деканские траппы

Во время мел-палеогенового вымирания активно извергались трапповые базальты[англ.] Деканских траппов в Индии. Извержения можно разделить на три фазы: две до мел-палеогеновой границы и одна после. Вторая фаза, которая произошла очень близко к границе, выбросила от 70 до 80% объёма всех остальных извержений в виде прерывистых импульсов, которые происходили с интервалом примерно в 100 000 лет[266][267]. Как предполагается, парниковые газы, такие как углекислый и сернистый газы, могли быть высвобождены этой вулканической активностью[268][269], став причиной температурных возмущений примерно на 3 °C или, возможно, до 7 °C, повлёкших за собой изменение климата[270]. Как и в случае с импактным событием, извержения могли также высвободить сульфатные аэрозоли, которые вызвали бы кислотные дожди и глобальное похолодание[271]. Однако из-за большой погрешности в датировке истинная роль извержений деканских траппов в мел-палеогеновом вымирании остаётся неясной[231][232][272].
До 2000 года аргументы о том, что вымирание вызвали извержения Деканских траппов, а не импактное событие, обычно связывались с мнением о том, что вымирание было постепенным. До открытия кратера Чикшулуб извержениями объяснялся глобальный иридиевый слой[268][273]; даже после открытия кратера часто считалось, что удар имел только региональное, а не глобальное влияние на вымирание[274]. В ответ Луис Альварес отверг вулканическую активность как объяснение иридиевого слоя и вымирания в целом[275]. С тех пор, однако, большинство исследователей заняло более умеренную позицию, согласно которой влияние импактного события считается первопричиной вымирания, а также признается, что Деканские траппы также могли сыграть свою роль. Сам Уолтер Альварес в конечном итоге признал, что Деканские траппы и другие экологические факторы, возможно, способствовали вымиранию в дополнение к импактному событию[276]. По некоторым оценкам, начало второй фазы извержений Деканских траппов наступило через 50 000 лет после импактного события[277]. В сочетании с математическим моделированием сейсмических волн эти оценки позволяют предположить, что импактное событие могло спровоцировать извержения Декановских траппов, увеличив проницаемость мантийного плюма, лежавшего под ними[278][279].
До сих пор нет однозначного мнения о том, были ли Деканские траппы основной причиной вымирания наравне с импактным событием. Сторонники существенной роли Деканских траппов считают, что климатическое воздействие выброса диоксида серы было на одном уровне с воздействием импактного события, а также отмечают роль паводкового базальтового вулканизма в других массовых вымираниях, таких как массовое пермское вымирание[280][281]. Они полагают, что импактное событие усугубило уже продолжающееся изменение климата, вызванное извержениями[282]. В то же время противники этой точки зрения утверждают, что воздействие траппов на мел-палеогеновое вымирание опровергается внезапным характером вымирания и существованием других импульсов активности Деканских траппов сопоставимой величины, по-видимому, не вызывали каких-либо массовых вымираний. Также они высказывали мнение, по которому причины различных массовых вымираний следует оценивать отдельно[283]. В 2020 году Альфио Кьяренца и его коллеги предположили, что Деканские траппы имели противоположный эффект: по их мнению, долгосрочное потепление, вызванное выбросами углекислого газа, могло смягчить импактную зиму[257].
Палеоценовые динозавры
Веерохвостые (кроновые) птицы считаются единственными динозаврами, достоверно пережившими мел-палеогеновое вымирание; тем не менее, известны некоторые спорные ископаемые остатки нептичьих динозавров, которые могут относиться к палеоцену[284]. В 2000 году Спенсер Лукас и коллеги сообщили об открытии правой бедренной кости гадрозаврида в бассейне Сан-Хуан в Нью-Мексико, назвав её свидетельством существования нептичьих динозавров в палеоцене. Породы, в которых была обнаружена кость, датируются возрастом в 64,8 ± 0,9 млн лет (ранний палеоцен)[285]. Если она не была повторно отложена в результате выветривания, то это означает, что некоторые популяции нептичьих динозавров могли существовать, по крайней мере, в течение полумиллиона лет после начала кайнозойской эры[286]. Также в пользу данной точки зрения свидетельствует присутствие остатков нептичьих динозавров в формации Хелл-Крик на высоте до 1,3 м над мел-палеогеновой границей, в отложениях, сформировавшихся через 40 000 лет после вымирания. Эти доказательства используются для поддержки мнения о постепенности мел-палеогенового вымирания[287]. Однако ископаемые остатки, на которых они основаны, по мнению многих других исследователей, могли быть переотложены, то есть вымыты из их первоначальных мест залегания, а затем снова захоронены в более молодых отложениях[288][289][290]. Кроме того, оценки их возраста иногда критикуются как ненадёжные[291].
Палеонтологическая реконструкция
Воссоздание внешнего облика нептичьих динозавров и представителей многих других вымерших таксонов в виде картин, скульптур, трёхмерных моделей и т. п. является задачей палеонтологической реконструкции[292]. Палеонтологическая реконструкция имеет высокое значение для формирования верного образа доисторического мира у широкой публики и отражения новых научных гипотез.
Принципы реконструкции в общих чертах одинаковы для всех групп позвоночных. Профессиональные палеохудожники создают свои работы на основе известного ископаемого материала — первым делом создаётся база в виде скелетной реконструкции; поскольку полные скелеты сохраняются довольно редко, обычно художнику приходится брать недостающие кости от разных образцов того же вида, либо другого, близкородственного ему. В дальнейшем на скелетную реконструкцию добавляются мягкие ткани, такие как мускулы и внутренние органы, благодаря чему животному придаётся его форма. На многих ранних реконструкциях динозавров у них почти отсутствовали мягкие ткани, в результате чего животные фактически представляли собой обтянутые кожей скелеты. Хотя мягкие ткани редко сохраняются в ископаемом состоянии, такой подход к реконструкции ныне считается неверным. Чтобы решить проблему недостатка материала, палеохудожники обращаются к тканям современных родственников нептичьих динозавров: так как птицы являются частью эволюционного дерева динозавров, особенности их анатомии и поведения имеют важную роль при создании реконструкции; однако поскольку птицы очень специализированы, иллюстраторы также используют в качестве образца анатомию нептичьих рептилий, таких как крокодилы и ящерицы[292].
В культуре

Сравнительная изученность как самих динозавров, так и драматической истории их вымирания в конце мелового периода, а также впечатляющие размеры многих из них, обусловили распространённость их образа в массовом сознании, сделав динозавров популярными «героями» многочисленных произведений. В культуре динозавры представлены прежде всего в качестве существ фантастической внешности и огромных размеров. Вхождение слова «динозавр» в бытовой язык отражает культурное значение этих животных: в переносном смысле слово «динозавр» часто используется для описания всего, что является непрактично большим, устаревшим или находящимся на грани исчезновения.
Общественный энтузиазм к динозаврам зарождался в викторианской Англии. В 1854 году, спустя три десятилетия после публикации первых научных описаний динозавров, в Лондоне был открыт парк Кристал-Палас[англ.], где до сих выставлены фигуры динозавров и других древних животных, реконструированных в соответствии с научными представлениями того времени. Фигуры из Кристал-Палас оказались настолько популярными, что вскоре появился сильный рынок их уменьшенных копий. Наличие большого числа хорошо сохранившихся ископаемых остатков, включённых в экспозиции крупнейших палеонтологических музеев мира, также весьма поспособствовало знаменитости этой группы. Неугасающая популярность динозавров, в свою очередь, привела к значительному государственному финансированию науки о динозаврах и часто стимулировала новые открытия. В США конкуренция между музеями за внимание общественности привела к Костяным войнам в 1877—1892 гг., в ходе которых двое соперничавших палеонтологов, Эдвард Дринкер Коп и Отниел Чарльз Марш, внесли огромный вклад в палеонтологию.
Популярность динозавров обеспечила им появление в литературе, кинематографе и других медиа. Динозавры были представлены в большом количестве художественных работ, начиная с краткого упоминания мегалозавра в романе Чарльза Диккенса «Холодный дом» (1852). Они имеют значительную роль в таких произведениях как романы «Путешествие к центру Земли» Жюля Верна (1864) и «Затерянный мир» сэра Артура Конан Дойла (1912), их экранизациях, в анимационном фильме «Динозавр Герти» (1914; первый фильм о динозаврах), фильмах «Кинг-Конг» (1934), «Годзилла» (1954), в романе-бестселлере Майкла Крайтона «Парк юрского периода» (1990) и снятой по его мотивам серии фильмов. Имеется ряд авторов научно-популярных работ о динозаврах, в том числе некоторые видные палеонтологи, которые могут пытаться использовать динозавров как способ рассказать читателям на их примере о науке в целом. В рекламе повсеместно встречаются динозавры; многочисленные компании упоминали динозавров в печатной или телевизионной рекламе для того, чтобы продвигать свои собственные продукты, а иногда для того, чтобы охарактеризовать своих конкурентов как медлительных, глупых или устаревших.
Геральдика

С точки зрения геральдической науки изображение нептичьего динозавра является естественным негеральдическим символом. Оно используется в символике тех местностей, где были найдены остатки динозавров, других древних животных, либо для привлечния внимания.
- На гербе и флаге Чебулинского района Кемеровской области присутствует изображение пситтакозавра, символизирующее открытия Александра Моссаковского.
- На гербе и флаге города Саки (Крым) присутствует стилизованное изображение динозавра.
- На гербе и флаге Чернышевского района Забайкальского края присутствует изображение пернатого динозавра кулиндадромея[293].
- На гербе Котласского района Архангельской области присутствует стилизованное изображение вымышленного динозавра, символизирующее открытия Владимира Прохоровича Амалицкого на его территории[294]. На самом деле Амалицкий занимался изучением пермских наземных позвоночных, живших ещё до появления динозавров.
Примечания
Литература
- Кэрролл Р. Палеонтология и эволюция позвоночных = Vertebrate Paleontology and Evolution / Под ред. акад. Л. П. Татаринова. — М.: Мир, 1993. — Т. 2. — 283 с. — ISBN 5-03-001819-0.
- Брусатти С. Время динозавров. Новая история древних ящеров = The Rise and Fall of the Dinosaurs: A New History of a Lost World / науч. ред. А. О. Аверьянов. — М.: Альпина нон-фикшн, 2019. — 358 с. — ISBN 978-5-91671-893-5.
- Нэйш Д.[англ.], Барретт П. Динозавры. 150 000 000 лет господства на Земле / науч. ред. А. О. Аверьянов, д-р биол. наук. — М.: Альпина нон-фикшн, 2018. — 223 с. — ISBN 978-5-91671-940-6.
- Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- Хоун Д. Хроники тираннозавра: Биология и эволюция самого известного хищника в мире = The Tyrannosaur Chronicles: The Biology of the Tyrant Dinosaurs / науч. ред. А. О. Аверьянов, д-р биол. наук. — М.: Альпина нон-фикшн, 2017. — 358 с. — ISBN 978-5-91671-744-0.
На английском языке
- Alvarez W. T. rex and the Crater of Doom (англ.). — Princeton, NJ: Princeton University Press, 1997. — 208 p. — ISBN 978-0-691-01630-6.
- Bakker R. T. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction (англ.). — New York: William Morrow and Company, 1986. — 481 p. — ISBN 978-0-688-04287-5.
- Benton M. J.[англ.]. Vertebrate palaeontology[англ.] (англ.). — 4th ed. — Wiley-Blackwell, 2015. — 480 p. — ISBN 978-1-118-40755-4. — ISBN 978-1-118-40684-7.
- The Complete Dinosaur (англ.) / M. K. Brett-Surman, T. R. Holtz Jr., J. O. Farlow, eds. — Bloomington, IN: Indiana University Press, 2012. — 1272 p. — (Life of the Past). — ISBN 978-0-253-00849-7.
- Brusatte S. L. Dinosaur Paleobiology (англ.) / ed. by M. J. Benton[англ.]. — University of California Press, 2012. — 336 p. — ISBN 978-0-520-25408-4.
- Encyclopedia of Dinosaurs (англ.) / ed. by Currie P. J., Padian K.[англ.]. — Academic Press, 1997. — 869 p. — ISBN 978-0-12-116810-6.
- Feathered Dragons: Studies on the Transition from Dinosaurs to Birds (англ.) / Currie P. J., Koppelhus E. B. Shugar M. A., Wright J. L., eds. — Bloomington, IN: Indiana University Press, 2004. — 361 p. — (Life of the Past). — ISBN 978-0-253-34373-4.
- Living Dinosaurs: The Evolutionary History of Modern Birds (англ.) / ed. by Gareth D. J.[англ.], Kaiser G.. — Chichester; Hoboken, NJ: Wiley-Blackwell, 2011. — 422 p. — ISBN 978-3-319-68008-8. — ISBN 978-3-319-68009-5.
- Fastovsky D. E., Weishampel D. B. Dinosaurs: A Concise Natural History (англ.). — 3rd ed. — Cambridge: Cambridge University Press, 2016. — 477 p. — ISBN 978-1-107-13537-6. — ISBN 978-1-316-50115-3.
- Fastovsky D. E., Weishampel D. B. The Evolution and Extinction of the Dinosaurs (англ.). — 2nd ed. — New York: Oxford University Press, 2005. — 485 p. — ISBN 978-0-521-81172-9. — ISBN 0-521-81172-4.
- Glut D. F.[англ.]. Dinosaurs: The Encyclopedia (англ.). — Jefferson, North Carolina: McFarland & Company, 1997. — 1076 p. — ISBN 0-89950-917-7.
- Gunther R. T., ed. Life and Letters of Edward Lhwyd (англ.) / Preface by A. E. Gunther (Reprint ed.). — London: Dawsons of Pall Mall, 1968. — Vol. XIV. — 576 p. — (Early Science in Oxford). — ISBN 978-0-7129-0292-2.
- Heilmann G.[англ.]. The Origin of Birds[англ.] (англ.). — London; New York: H. F. & G. Witherby[англ.]; D. Appleton & Company[англ.], 1926. — 209 p.
- Lessem D.[англ.], Glut D. F.[англ.]. The Dinosaur Society's Dinosaur Encyclopedia (англ.) / Illustrations by T. L. Ford; scientific advisors, P. Dodson[англ.], et al.. — New York: Random House, 1993. — 533 p. — ISBN 978-0-679-41770-5.
- Lhuyd E. Lithophylacii Britannici ichnographia (англ.). — London: Ex Officina M.C, 1699.
- Mayr G.[англ.]. Paleogene Fossil Birds (англ.). — Berlin: Springer-Verlag, 2009. — 262 p. — ISBN 978-3-540-89627-2.
- Molina-Pérez R., Larramendi A. Dinosaur Facts and Figures: The Theropods and Other Dinosauriformes (англ.) / Illustrations by A. Atuchin and S. Mazzei. — Princeton, New Jersey: Princeton University Press, 2019. — 288 p. — ISBN 978-0-691-18031-1.
- Norell M. A.[англ.], Gaffney E. S.[англ.], Dingus L. W. Discovering Dinosaurs: Evolution, Extinction, and the Lessons of Prehistory (англ.). — Revised ed. — Berkeley: University of California Press, 2000. — 239 p. — ISBN 978-0-520-22501-5.
- The Origin of Birds and the Evolution of Flight (англ.) / ed. by K. Padian[англ.]. — San Francisco, CA: California Academy of Sciences, 1986. — Vol. 8. — 98 p. — (Memoirs of the California Academy of Sciences). — ISBN 978-0-940228-14-6.
- Parsons K. M. Drawing out Leviathan: Dinosaurs and the Science Wars (англ.). — Bloomington, IN: Indiana University Press, 2001. — 210 p. — (Life in the Past). — ISBN 978-0-253-33937-9.
- Paul G. S.[англ.], ed. The Scientific American Book of Dinosaurs (англ.). — 1-st ed. — New York: St. Martin's Press[англ.], 2000. — 432 p. — ISBN 978-0-312-26226-6.
- Paul G. S.[англ.]. Predatory Dinosaurs of the World: A Complete Illustrated Guide (англ.). — New York: Simon & Schuster, 1988. — 464 p. — ISBN 978-0-671-61946-6.
- Paul G. S.[англ.]. The Princeton Field Guide to Dinosaurs[англ.] (англ.). — Princeton: Princeton University Press, 2016. — 360 p. — ISBN 978-0-691-16766-4.
- Plot R.[англ.]. The Natural History of Oxford-shire: Being an Essay toward the Natural History of England (англ.). — 1677.
- Schutt B. Cannibalism: A Perfectly Natural History (англ.). — New York: Algonquin Books, 2017. — 352 p. — ISBN 1616204621. — ISBN 978-1616204624.
- The Dinosauria[англ.] (англ.) / D. B. Weishampel, P. Dodson, H. Osmólska, eds. — 2nd ed. — Berkeley: University of California Press, 2004. — 880 p. — ISBN 978-0-520-25408-4.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K.[англ.], Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.
- Holtz Jr. T. R. Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages (англ.) / Illustrated by L. V. Rey. — New York: Random House, 2007. — 432 p. — ISBN 978-0-375-82419-7.
Ссылки


 Птицы - это динозавры? Рассказывает Павел Скучас
Птицы - это динозавры? Рассказывает Павел Скучас- The Theropod Database — база данных мезозойских ящеротазовых (англ.).
- Майкл Грешко (2020-10-29). "Реальные динозавры: какими они были на самом деле". National Geographic. Дата обращения: 6 апреля 2021.
- Michael Greshko. See how we're reimagining dinosaurs in today's 'golden age' of paleontology (англ.). National Geographic. Дата обращения: 6 апреля 2021. — Оригинал публикации с бо́льшим количеством иллюстраций и анимациями.
- Панорамные виды Палеонтологического музея имени Ю. А. Орлова (Москва).
- Панорамные виды Национального музея естественной истории (Вашингтон) (англ.).
- 3D-модель Палеонтологического музея в Париже (англ.).
- TaxonSearch — база данных филогенетических определений (англ.).